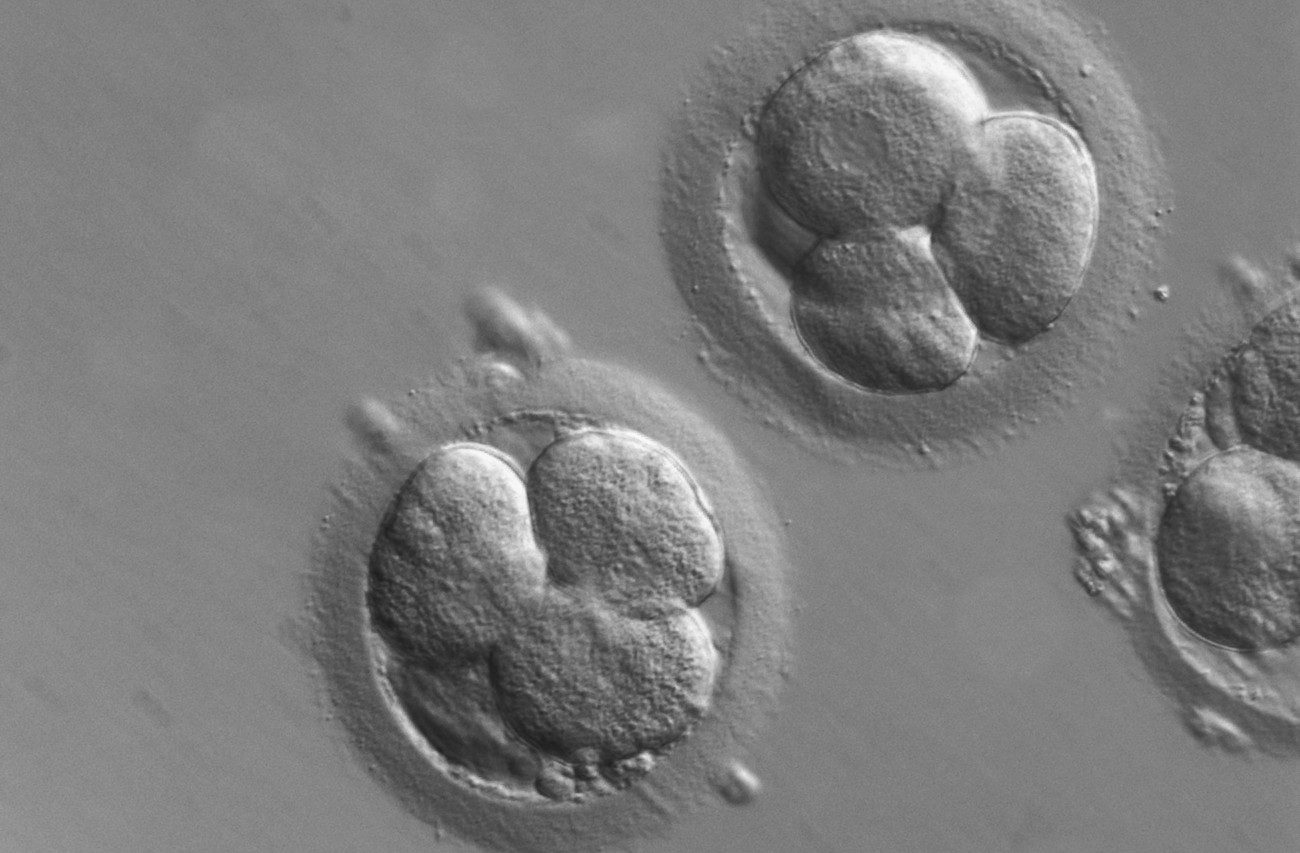
In the debate about the moral standing of human embryos, some defenders of embryo-destructive research have claimed that human embryos are not human beings until implantation (i.e., when the embryo attaches to the uterus, approximately six days after fertilization), and others have claimed that they are not human beings until gastrulation (i.e., when the possibility of twinning no longer exists and the primitive neural streak first appears, approximately 14 days after fertilization). These claims have been repeated by policymakers, scientists, and bioethicists alike, yet they fly in the face of the embryological evidence. Seeing why will put the embryo research debate on a more solid biological footing.
Over the past few years, Utah Senator Orrin Hatch has pushed aggressively for federal funding of embryo-destructive research. When it comes to abortion, Senator Hatch votes consistently pro-life; he believes we have a moral obligation to protect developing human beings. But he also believes that embryos produced outside of a woman’s body, whether by cloning or in vitro fertilization, are not human beings unless or until they are implanted in a uterus. “At the core of my support for regenerative medicine research,” he declared in 2002, “is my belief that human life requires and begins in a mother’s nurturing womb.”
More recently, William Neaves, president of the Stowers Institute for Medical Research in Kansas City, has similarly claimed in public hearings that the embryo does not become a human being until implantation. According to Neaves, not until the embryo receives external, maternal signals at implantation is it able to establish the basic body plan of the human, and only then does it become a self-directing human organism. According to Neaves, these signaling factors somehow transform what was hitherto a mere bundle of cells into a unitary organism.
In reply to Hatch, Neaves, and others who make this argument, the first point to notice is that the standard embryology texts locate the beginning of the human individual at fertilization, not at implantation. See, for example, William J. Larsen, Human Embryology, 3rd ed. (2001); Keith Moore and T.V.N. Persaud, The Developing Human, Clinically Oriented Embryology, 7th ed. (2003); and Ronan O’Rahilly and Fabiola Mueller, Human Embryology and Teratology, 3rd ed. (2000). Most people who point to implantation as the beginning of an individual human life — Senator Hatch is a prime example — offer not the slightest bit of evidence to support their claim, relying instead on an alleged intuition. But since such intuitions can be matched by contrary intuitions, and since the alleged intuitions of Hatch and others contradict the evidence supplied by embryological science, they have no evidential weight whatsoever.
Neaves does offer an actual argument, but it is severely flawed. He claims that at implantation maternal signaling factors transform a bundle of cells into a human organism. But there is much dispute about whether any such maternal signaling actually occurs. As Hans-Werner Denker observes, it was once assumed that in mammals, in contrast to amphibians and birds, polarity in the early embryo depends upon some external signal, since no clear indications of bilateral symmetry had been found in oocytes, zygotes, or early blastocysts. But this view has been revised in light of emerging evidence: “[I]ndications have been found that in mammals the axis of bilateral symmetry is indeed determined (although at first in a labile way) by sperm penetration, as in amphibians. Bilateral symmetry can already be detected in the early blastocyst and is not dependent on implantation.”
Denker refers specifically to the work of Magdalena Zernicka-Goetz and her colleagues at Cambridge and that of R. L. Gardner at Oxford, which shows that polarity exists even at the two-cell stage. Davor Solter and Takashi Hiiragi of the Max Planck Institute for Immunobiology in Freiburg dispute these results, arguing that in the early embryo (prior to compaction and differentiation into inner cell mass and trophoblast) external factors determine the fate of each cell, rather than an internal polarity. As Gretchen Vogel reported in 2005 in Science magazine, embryologists are “polarized over early cell fate determination.” It is no longer taken as certain that the bilateral polarity of the embryo does not occur in the very first cleavages.
Moreover — and more importantly — even if it is the case that polarity does not emerge until a maternal signal is received at implantation, that would not provide any evidence at all that such a signal transformed a bundle of cells into a unitary, multicellular human organism. Rather, just as the lungs begin to breathe at birth only in response to certain external stimuli, so it would make sense that differentiation into the rudiments of the distinct body parts (basic bilateral polarity) would begin only in response to some external stimuli. And this is exactly how such signals speculated to occur (perhaps) in mammalian embryos are interpreted by the embryology texts that mention them. Thus, Neaves not only treats uncertain data as definitive, but — more to the point — his claim fails to hold up even if, for the sake of argument, one grants his assumptions.
The last point in reply to the claim that the human being is not generated until implantation is the most important one: there is complex and coordinated development from day 1 to day 6, much of it plainly oriented to preparing the embryo for the implantation process, as well as for processes that will occur only after that. The proposition that the human organism does not come to be until implantation (day 6) offers no explanation for this regular and ordered development.
On day 3 or 4 compaction occurs, which is the process in which the cells change their shapes and align themselves closely together. And compaction is the first step toward cavitation — the process (at day 4) in which an inner cavity is formed within the embryo and the embryo differentiates itself into the inner cell mass (which will later develop into the body of the mature organism) and the trophoblast (which will later develop into the placenta, a temporary organ of the embryo, equivalent to other temporary parts of the body, like baby teeth). On day 5 or 6, as the embryo enters the uterus, it “hatches” from the zona pellucida — the membrane enveloping the ovum that the sperm had to penetrate for fertilization to occur — preparing to begin implantation. At the same time, the trophoblast cells secrete an enzyme which erodes the epithelial lining of the uterus and creates an implantation site for the embryo.
In addition, the trophoblast itself becomes differentiated (about day 3 or 4) into various levels (cytotrophoblast and syncytiotrophoblast) in preparation for developing the vital contacts with the mother’s blood system (the embryo will circulate its own blood but will exchange oxygen and wastes with the mother’s blood, first through connecting microvilli, and eventually through the umbilical cord, developed from the trophoblast). Around the same time, the trophoblast produces immunosuppressive factors signaling the mother’s system to accept the embryo rather than attack it as a foreign substance. In order for the embryo as a whole to survive, this complex series of activities must occur in a timely, ordered sequence and with predictable regularity. Clearly, these activities — compaction, cavitation, and implantation itself — are organized processes performed by the embryo as an organismal whole.
The test of whether a group of cells constitutes a single organism is whether they form a stable body and function as parts of a whole, self-developing, adaptive unit. Compaction, cavitation, the changes occurring earlier to facilitate these activities, and implantation — all of these activities are clear cases of the cells acting in a coordinated manner for the sake of a self-developing and adaptive whole. In other words, such activities are ordered to the survival and maturation of the whole, existing embryo. This fact shows that the unity of the blastomeres (the cells of the early embryo) is substantial rather than incidental; the blastomeres are integrated parts of a functional whole, not separate parts that lead to the creation of a whole. This is compelling evidence that what exists from day 1 to day 6 is not a mere aggregate of cells but a multi-cellular organism.
Of course, one might object that even if there is an organism from day 1 to day 6, perhaps it is not the same as the organism after day 6. Perhaps implantation and its concomitant events produce a substantial change, the generation of a new organism. In reality, however, the direction of the growth between day 1 and day 6, on the one hand, and from day 6 onward, on the other hand, is the same. That is, the sequence of steps in the embryo from day 1 to day 6 is necessary and preparatory for what occurs afterward, and is a unitary trajectory of development. It is unlike, for example, the separate sequences of events undergone by the sperm and the ovum, respectively, before fertilization. Gametes (sperm and ovum) are oriented to joining with each other, actions that are performed not by them as a single unit, but by the maternal and paternal organisms (i.e., the mother and father). The sperm and the ovum (prior to fusing) are distinct biological parts of the distinct parent organisms (even though in coitus a type of organic union is effected between the male and female organisms). By contrast, the human embryo’s cells (from day 1 onward) form a stable body and work together to produce a single direction of growth, which is toward the maturation of the human organism.
The actions of the embryo from day 1 to day 6 are clearly part of a unitary development toward human maturation. None of the events occurring in the embryo could reasonably be interpreted as creating a new and distinct direction. Implantation does not change the nature (kind of being) of the embryo; it is an event in the unfolding life of a whole human organism, not the initiation of an entirely new organism.
Another attempt to locate the beginning of the human being after fertilization is based on the rare phenomenon of monozygotic twinning and the even rarer phenomenon of fusion. Monozygotic twinning occurs when embryonic division results in two whole embryos. Apparently, fusion can also occur in humans: that is, two embryos can fuse to become one embryo. Such twinning and fusion are possible up to approximately day 14, with the appearance of the primitive streak, the visible precursor of the spinal cord and brain. Some argue that the possibility of twinning or fusion shows that prior to day 14 the embryo is not an individual; the individual who is clearly present at more mature stages of development has not yet come to be. Raymond Devettere expresses the argument clearly:
If we say a zygote is one of us, then we are also saying that one of us can become two of us, and that two of us can become one of us. This makes no sense. The possibility of the zygotes’ splitting or fusing suggests the zygote is not yet what we mean by one of us.
The puzzle, some seem to suppose, is that if we trace someone’s life back to adolescence, then to infancy, then to fetal existence, and then back to the embryonic stage, it does not seem as though the individual’s life traces back to fertilization. The tracing, in some cases, seems to stop at twinning or fusion.
But there is no puzzle here. Although twinning and fusion raise interesting questions about the details of early embryonic life, the argument that an individual life is not yet present is simply fallacious. Rather, in twinning, either the first embryo dies and gives rise to two others, or the first embryo continues to live and a second embryo is generated upon the splitting of the first one. We think the latter alternative is more likely, that twinning is (like induced cloning) a type of asexual reproduction in which the second embryo is reproduced asexually. For although monozygotic twinning can occur at the two-cell stage, most monozygotic twinning (at least two-thirds) occurs between days 5 and 9. In those cases, the growth trajectory of the original embryo continues, though the separation of some of the cells from the inner cell mass generates another embryo, with a distinct development trajectory. (If the splitting occurs after day 9, the embryos may share some of their permanent organs, resulting in conjoined twins, which are two distinct organisms that possess some degree of organic union.) The possibility of embryo fusion also poses no difficulty for the individuality of the embryo from day 1. If fusion in fact occurs, one embryo is absorbed into the system of another.
In a recent issue of Commonweal, Cathleen Kaveny attacks the position that an individual human life begins at fertilization, as it was articulated years ago by Germain Grisez:
Grisez’s attempt to preserve the claim that individuated human life begins at fertilization [by arguing, as we have, that twinning is a type of asexual reproduction] sacrifices too much of what we know about human nature — both from a Christian perspective and a scientific one. After all, human beings reproduce sexually, not asexually. Humans are mortal; they die and their bodies disintegrate. They don’t split neatly into two twins with no loss, cost, or remainder (as in twinning), nor do they merge fluidly into one another (as in combination).
Here we have a parade of assertions presented as if they constituted an argument. To assert that human beings never reproduce asexually is no argument at all, but simply a denial of the opposite position. Asexual reproduction is not the norm in humans, but embryological evidence shows that human beings in the early embryonic stage possess the capacity for asexual reproduction — a capacity that requires special environmental circumstances to induce it. Likewise, to say that when humans die their bodies always disintegrate, that their cells could never be absorbed by another, is just presupposing the falsity of the contrary position, rather than coming to terms with the scientific facts on which it is based.
When Kaveny asserts that our position “sacrifices too much of what we know about human nature — both from a Christian perspective and a scientific one,” she gives no indication of how it is supposed to conflict with Christian teaching. However, what we know about human nature from a Christian standpoint is fully compatible with the position set out above: namely, that a human individual is present even while the possibilities of fusion and twinning remain. Let us assume that each human being receives a rational soul when he comes to be. Just how he comes to be (whether immediately at fertilization or a few days later upon a splitting off of some of the first embryo’s cells) is irrelevant to that belief. And from a scientific standpoint, the position we have set out provides an intelligible explanation of the coordinated development of the embryo from day 1 up to twinning, should twinning occur, whereas the denial that there is an individual human organism prior to that point leaves such detailed coordination inexplicable.
Viewed biologically, the occurrence of monozygotic twinning and the possibility of fusion fail to show that in the first fourteen days the cells within the embryo constituted only an incidental mass. Just as the division of a single, whole flatworm into two whole flatworms does not show that prior to that division the flatworm was not a unitary individual, just so with the human embryo that twins. Parts of a flatworm have the potential to become a whole flatworm when isolated from the present whole of which they are part. Likewise, at the early stages of an embryo’s development, the degree of cellular specialization has not progressed very far (even if the process of orderly cellular activity is underway from the beginning), which means the embryo’s cells or groups of cells can become whole organisms if they are divided and have an appropriate environment after the division. But that does not show that prior to such an extrinsic division the embryo is a mere mass of cells rather than a single, complex, actively developing human organism.
There is additional, decisive evidence for this point: if the individual cells within the embryo before twinning were each independent of the others, there would be no reason why each would not regularly develop on its own. But as we know, these allegedly independent, non-communicating cells actually function together to develop into a single, more mature member of the human species. This fact shows that interaction is taking place between the cells from the very beginning (even within the zona pellucida, before implantation), restraining them from individually developing as whole organisms and directing each of them to function as a part of a single, whole organism identical with the zygote. This means that prior to an extrinsic division of the embryo’s cells resulting in the existence of a twin, these cells together constitute a single organism. And prior to the (even rarer) event of fusion, the twin embryos that fuse are distinct, whole organisms exhibiting active development.
Science has not solved every mystery of early human development. But human embryology has advanced sufficiently to enable us to dismiss certain fallacies about when a new human life comes to be. We do not doubt the good faith of those who believe that individual life begins at implantation or after the powers of twinning and fusion have passed. But arguments advanced to support these beliefs collapse under scrutiny. We must not let the desire to use human embryos in research obscure our grasp of what those embryos truly are from day 1: namely, nascent members of the human species, worthy of that fundamental respect and protection that justice demands for every member of the human family.
Exhausted by science and tech debates that go nowhere?